| |
 |
|
|
|
ANT
ETOLOGIA
|
|
|
 |
 |
|
|
 |
Il
compito dell'agente è quello di ritornare al punto di partenza
da qualsiasi punto dello spazio si trovi (homing).
Cerchiamo ora di definire il concetto di homing nella sua versione più
semplice, ovvero quella che non coinvolge l'uso di mappe cognitive. Per
fare ciò ci riferiremo al comportamento di homing che si osserva
presso le formiche del deserto, Cataglyphis fortis. In questa pagina
verranno illustrati gli esperimenti etologici più significativi
in proposito.
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
L'homing
nelle formiche del deserto
|
|
|
|
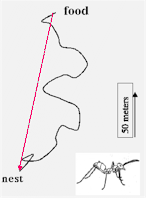
Quando
le formiche, o le api, lasciano il nido per cercare il cibo, compiono delle
traiettorie tortuose. Le formiche del deserto possono arrivare anche a 100
metri dal nido. Quando trovano il cibo mostrano un comportamento caratteristico:
si voltano e si dirigono direttamente verso il
nido lungo la direzione che collega il nido al luogo dove si trovano attualmente.
|
|
|
|
|
|
|
|
| |
 |
| |
|
| |
| |
|
Si muovono rapidamente
(circa 15 metri al minuto) attraverso il terreno del deserto privo di
punti di riferimento, finché, a pochi metri dal nido, mostrano
un comportamento di ricerca di punti di riferimento, o landmarks, familiari
(GALLISTEL, 1990).
Secondo l'autore le spiegazioni di tale tipo di comportamento possono
essere due:
1.
calcolano e mantengono la direzione verso casa attraverso un meccanismo
di piloting, cioè di stima della posizione attraverso il riconoscimento
di landmarks familiari (nel qual caso dovrebbero avere una mappa del
terreno che si estende per circa 100 metri dal nido);
2.
computano continuamente la loro posizione relativa al nido (vettore
direzione) per tutto il tempo (un'ora e più) durante il quale conducono
la loro ricerca e usano questo vettore per calcolare la correzione nella
traiettoria per intraprendere la via di casa.
Le formiche del deserto
usano il secondo metodo (direzione computata) per calcolare la
rotta verso il nido e mantenere tale rotta attraverso l'orientamento con
il sole e la stima di quanto lontano sia il nido, in modo da innescare
il meccanismo di piloting (ricerca di landmarks familiari) al momento
giusto (in prossimità del nido).
Infatti esse non hanno una mappa del territorio attorno al nido.
Si considerino, infatti,i seguenti esperimenti.
|
| |
|
|
|
Figura 1.
Homing nelle formiche del deserto. A: percorso di homing di una formica.
S: punto del rilascio. N*: nido fittizio. O: svolta indicante il fatto
che le formiche cominciano a cercare il nido (piloting). B: punti che
stanno per la posizione di dieci formiche rilevata ogni 10 secondi dal
momento che incominciano il percorso di ritorno dal punto di rilascio.
C: posizione dei 10 formiche rilevata ogni 10 secondi, dopo 15 minuti
che hanno incominciato la ricerca del nido. Da
GALLISTEL, 1990.
|
| |
| |
|
[1]
WEHNER e FLATT (1972).
Le formiche catturate non appena uscivano dal nido e rilasciate
a tre metri di distanza da esso non mostravano in nessun modo di conoscere
la direzione del nido. Solo il 57 per cento di esse riuscì a tornare
al nido in meno di cinque minuti. Il tempo medio di 2,18 minuti impiegato
da questo 57 per cento è superiore di dieci volte il tempo impiegato
da una formica, che è già sulla via del ritorno, per percorrere
la stessa distanza.
|
| |
| [2]
WEHNER e SRINIVASAN (1981). Il
cibo è piazzato a 20 e 40 metri dal nido. A circa 600 metri dal nido
e dal cibo gli autori disegnarono un reticolo di linee a circa un metro
l'una dall'altra. Le formiche erano catturate quando avevano trovato il
cibo e si apprestavano a muoversi verso il nido. Venivano poi rilasciate
presso la griglia disegnata a 600 metri di distanza. Il punto di rilascio
è indicato dal quadratino vuoto etichettato con S in figura 1.1,
mentre il punto dove avrebbe dovuto trovarsi il nido se la griglia fosse
sovrapposta al territorio da cui le formiche sono state rimosse è
indicato con N*. Come previsto, le formiche non si accorsero dello spostamento
e, una volta liberate, intrapresero il percorso diretto verso il posto dove
si sarebbe dovuto trovare il nido. Il percorso lineare delle formiche terminava
con una svolta brusca (O). Il punto di svolta indicava il punto in cui esse
incominciavano a cercare in nido, o meglio la loro stima di dove avrebbe
dovuto essere. Le formiche cominciano a cercare il nido, mostrando un comportamento
di ricerca di punti di riferimento familiari, o piloting. Tale comportamento
mostra due importanti proprietà: rimane centrata sulla stima originale
del nido e nel punto stimato gli attraversamenti sono maggiori (figura 1
C). |
|
| |
| [3]
(stesso esperimento) Si cattura la formica
mentre sta cercando il nido nella posizione stimata e la si rilascia circa
10 metri lontano: se la formica prendesse dei punti di riferimento (landmarks)
in prossimità del punto di inizio della ricerca e successivamente
usasse il piloting per riferirsi ad essi, allora dovrebbe continuare a condurre
una ricerca centrata sul punto iniziale, dovrebbe cioè ritornare
alla posizione stimata iniziale (al punto di cattura); invece la nuova ricerca
è centrata sul punto di rilascio e non sul punto di cattura. Il vettore
fra il punto di cattura e il punto di rilascio è uguale al vettore
fra il centro della ricerca precedente alla cattura e il centro della ricerca
successiva. |
| |
| [4]
SANTCHI (1913).
Una formica procedeva con il sole alla sua destra. L'autore oscurò
la sua visione diretta del sole e mise uno specchio alla sinistra della
formica in modo che essa potesse vedere l'immagine riflessa del sole sullo
specchio dalla parte opposta a quella reale. La formica tornò indietro
e intraprese una traiettoria nella direzione opposta. SANTCHI osservò
che mutando l'angolazione dello specchio la formica cambiava l'angolazione
della sua traiettoria in modo da mantenere costante l'angolo relativo dell'immagine
dell'azimut del sole. |
| |
| [5]
JANDER (1957). Quando
un viaggio rettilineo di una formica viene interrotto dall'imprigionamento
della medesima in una scatola che le nasconde la vista del sole anche per
18 ore, la direzione che segue dopo l'imprigionamento non devia in media
dalla direzione seguita prima dell'imprigionamento. Essendo la velocità
angolare del sole di 15 gradi all'ora la formica, dopo 10 ore per esempio,
ha dovuto estrapolare il cambiamento di 150 gradi nell'azimut del sole.
|
| |
| [6]
RENNER (1959). Addestrò
le api in un campo aperto a Long Island a volare in un posto dove c'era
il cibo situato a nord-ovest dell'alveare (315°). Una notte trasferì
l'alveare in California, a Davis, in un campo aperto simile a quello di
Long Island. Egli piazzò dei punti dove le api avrebbero trovato
il cibo a 45° di intervallo attorno all'alveare in California con un
raggio uguale alla distanza del cibo a Long Island. L'ipotesi era che, siccome
la differenza di fuso orario fra i due posti è di circa tre ore,
l'orologio endogeno delle api si sarebbe trovato tre ore avanti. Esse avrebbero
commesso un errore proporzionale nella stima della direzione del cibo rispetto
al sole. Infatti, essendo una caratteristica delle api andare verso il cibo
alla stessa ora, e percorrendo il sole 15° all'ora in media, le api
avrebbero dovuto sbagliare la stima di 45 gradi in difetto. In effetti la
maggior parte delle api a Davis si diresse verso il cibo situato a 270°
(315° - 45°) in linea con l'ipotesi. |
| |
|
Partendo
da tali evidenze sperimentali vediamo di riassumere le caratteristiche
dell'homing nella sua versione più semplice.
|
| |
|
In base ai risultati
di [1] e [2] le formiche non sanno dove sono finché non vi ci si
trovano e ciò suggerisce che sanno dove sono attraverso il dead
reckoning (GALLISTEL, 1990). Il dead reckoning
genera la più semplice delle relazioni geometriche fra due punti
sulla superficie terrestre, il punto di partenza e la posizione corrente.
Secondo l'autore questo avverrebbe attraverso la capacità degli
insetti di integrare la velocità rispetto al tempo.
Gli esperimenti [1],
[2] e [3] dimostrerebbero inoltre che la stima del punto di partenza
non avviene attraverso l'uso di una mappa cognitiva. Il dead reckoning,
ovvero la stima del vettore fra il punto attuale ed il punto di partenza,
è l'unica funzione implicata.
In base all'esperimento [4] il procedimento di stima del punto di partenza
avviene prendendo come punto di riferimento il sole, o meglio l'azimut
del sole, cioè la proiezione della sua direzione sul piano bidimensionale
su cui si muove la formica.
Il sole non è un punto di riferimento fisso: infatti il cambiamento
nell'azimut del sole è in media di circa 15° all'ora. Pertanto,
pur potendolo considerare un punto di riferimento fisso in senso relativo,
in quanto il suo movimento è estremamente più lento del
movimento dell'insetto, non lo è in senso assoluto. In altre parole
se la formica o l'ape mantenessero costante l'allineamento con l'azimut
del sole, il loro percorso sarebbe curvilineo, in quanto l'azimut del
sole si sposta in funzione del tempo.
Gli esperimenti [5]
e [6] dimostrano che gli insetti sono in grado di correggere l'angolo
di allineamento con il sole in modo da annullare (o minimizzare) gli effetti
del cambiamento nell'azimut del sole. In qualche modo essi devono
avere una funzione effemeride interna, che computa il cambiamento nella
posizione dell'azimut del sole in funzione del tempo in base all'orologio
interno in fase con il ciclo cicardiano (GALLISTEL,
1990).
L'esperimento [6]
dimostrerebbe proprio questa caratteristica.
|
| |
L'osservazione
del comportamento dell'agente biologico ci costringe a fare delle assunzioni
e VINCOLA la progettazione dell'agente artificiale
|
| |
|
|
|
©
2011 Emernet
|